Introduction
Introduction to Pulmonary Fibrosis
Pulmonary Fibrosis (PF) describes a dissimilar group of lung disorders characterized by progressive and irreversible destruction of lung architecture giving rise to scar formation[1]. The pathologic changes that characterize idiopathic pulmonary fibrosis varies by location and age of the lesions, but predominantly found in the peripheral subleural parenchyma. Fibrotic zones with associated honeycombing alternate with healthy parts of lung tissue within fibrotic zones, foci of actively proliferating fibroblasts and myofibroblasts are presented (Fig. 1).
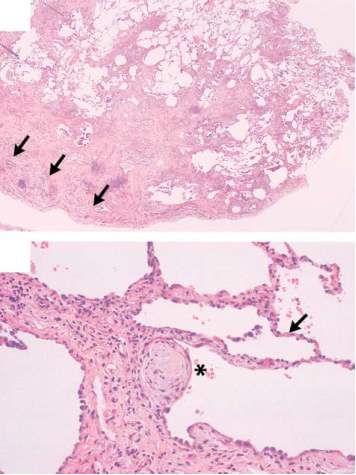
These focal zones of fibroblast proliferation (“fibroblast foci”) occur at sites of recent alveolar injury. The active cellular response at these areas is similar to healing patterns observed in skin and other tissues. As the inflammatory responses leads to leukocyte recruitment and angiogenesis, the perseverance of inflammation through apoptotic and phagocytic pathways restores normal tissue architecture. However, chronic exposure to irritants in the form of allergens, toxic chemicals (e.g. chemotherapy agents) and radiation results in dysregulated wound healing response, constant inflammation and ultimately fibrosis.
Scar formation disrupts normal physiologic lung function (e.g., breathing) leading to organ breakdown, interruption of gas exchange from alveoli into the blood and finally death due to respiratory failure. Pulmonary injury and infection cause destruction of lung architecture resulting in acute inflammation, a wound healing response [1], [2].
The interstitial inflammation of idiopathic pulmonary fibrosis is mild and generally associated with fibrosis. Thus, idiopathic pulmonary fibrosis is characterized by sequential acute lung injury that results in a progressive accumulation of fixed fibrosis with architectural distortion. As mentioned, fibroblast foci are the hallmark of idiopathic pulmonary fibrosis. These sites are characterized by extreme replication of activated fibroblast cells and dynamic deposition of fresh extracellular matrix. Cytokines normally associated with wound healing, such as transforming growth factor-b (TGF-β) and platelet derived growth factor (PDGF), are expressed at sites of injury. The driving force for this focal acute lung injury and wound repair is unknown and more current hypothesis for the pathogenesis of idiopathic pulmonary fibrosis suggests that a still-unidentified stimulus produces repeated episodes of acute lung injury Wound healing at these sites of injury ultimately leads to fibrosis, with loss of lung function. Therefore, one effective therapeutic strategy might be to modify fibroblast replication and matrix deposition.
Fibroblasts
In a healthy lung, small population of fibroblast cells reside in normal alveolar septae, these fibroblast cells are contractile yet do not express α- Smooth muscle actin [2], [3]. Myofibroblast over activity has been directly correlated with progression of lung fibrosis and deposition of collagen type I (Figure 2).
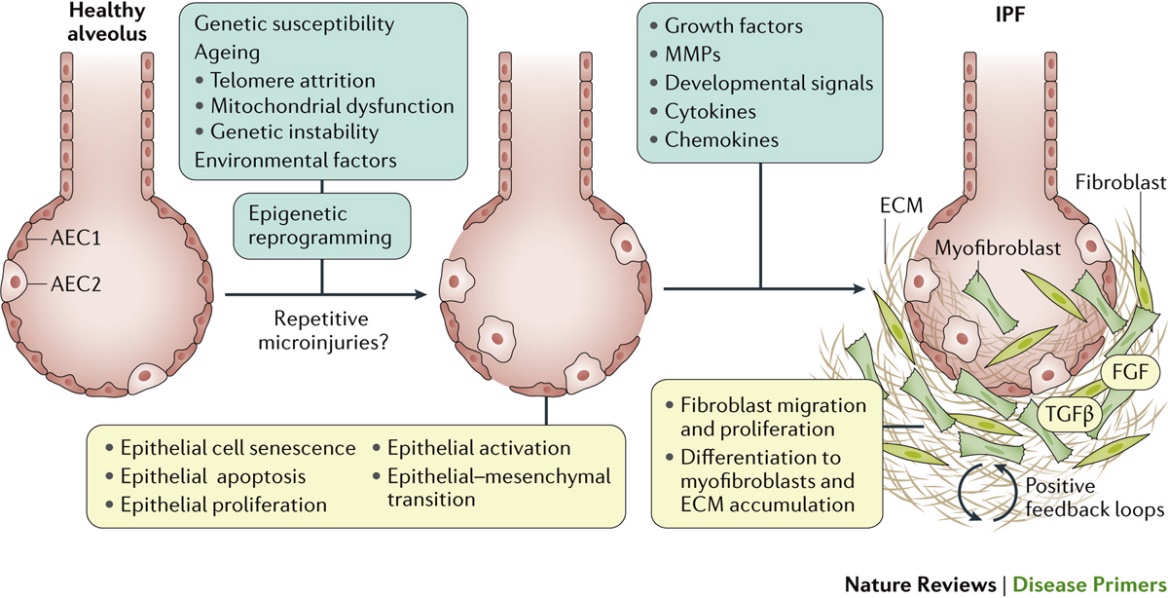
Transforming Growth Factor b1 (TGFb1) activates residing local fibroblasts, other resident cells, and blood-circulating precursor fibroblast cells into myofibroblasts. Myofibroblast activation is characterized by de novo expression of a-smooth muscle actin (α-SMA) in stress fibers, this cellular transfomation results in increases in cellular contractility hence building up mechanical tension in the tissue[4], [5]. The presence of myofibroblasts is important for restoring the integrity of damaged tissues by producing extra cellular matrix and forming a collagenous scar. The scar protects injured organs from further damage upon mechanical challenge, such as the heart after myocardial infarction, skin after trauma, and tendon, bone, and cartilage after fracture or rupture. However, in many conditions, transient and controlled myofibroblasts express resistance to their physiological clearance through apoptosis. In fibrosis, the excessive and detrimental myofibroblast activities turn beneficial tissue repair into devastating tissue deformation. This accumulation of a fibrotic scar within the otherwise sparsely inhabited interstitium results in both decreased lung compliance (i.e., increasing stiffness) and loss of gas exchange, ultimately rendering patients unable to breathe [1–4].
Many myofibroblast precursors are mesenchymal and locally available, including fibroblasts and mesenchymal progenitor cells that reside in the connective tissue architecture of all organs. Plus, myofibroblasts are generated from circulating progenitors, such as fibrocytes of hematopoietic origin and bone marrow–derived mesenchymal stem cells[1], [2] and [6]. However, when it comes to lung fibroblast cells studies were conducted to find out the origin if these cells based on observations of its cytoskeletal phenotype, tissue localization, and in vitro studies. Based on induction kinetic and expression of α-SMA perivascular and peribronchiolar adventitial fibroblasts are suggested as precursors[6], [7]. Additionally, circulating fibrocytes which express biomarkers such as CD45, CD34, collagen I, and CXCR4 have been reported to migrate to sites of tissue injury and differentiate into myofibroblasts. Studies that used BM chimeric mice, in which the donated BM cells can be tracked, as well as human transplant studies, showed that BM-derived progenitors can produce lung fibroblasts[7]. However, the ability of BM-derived fibroblasts to differentiate to myofibroblasts still could not be verified in some studies. Therefore, differentiation of stem cells into myofibroblasts was not confirmed by other studies. Hence, to date it is still unclear that lung myofibroblasts are originated from which cell populations and what is their relative contributions to the development of lung fibrosis. It is unlikely that this question can ever be answered in a definitive way. It is plausible that, the type of injury, the local environment, and the progression of the disease will produce different myofibroblast populations.
General characteristic of myofibroblasts, no matter the origin conditions and precursors, is the production of a collagen ECM and its remodeling by exerting high contractile forces[4], [7]. Despite their phenotypically resemble to smooth muscle cells (SMCs), the irreversible outcome of myofibroblast contraction suggests that it is somewhat differently regulated.
In SMCs, increased levels of cytosolic Ca2+ enhance the activity of the myosin light chain (MLC) kinase through the Calcium/calmodulin pathway[8], [9], which in turn MLC phosphatase activity determines the force development. However, in fibroblastic cells, the MLC phosphatase appears to be the key regulatory enzyme[9]. Fibroblast contraction is achieved by inactivation of the MLC phosphatase through the Rho-(associated) kinase (ROCK or ROK), a downstream target of the small GTPase RhoA, in contrast to regulation by Ca2+ cytosolic presence. Unlike SMC contraction, RhoA activity regulates fibroblast contraction and sufficiently high cytosolic calcium levels affects continued MLC activity[10]. Previous studies on role of myofibroblasts in wound healing exhibited RhoA as the chief regulator of myofibroblast contractility. Inhibition of MLC phosphatase using the phosphatase inhibitor calyculin was shown to be sufficient to induce strip contraction, whereas increasing cytosolic Ca2+ by membrane depolarization had little effect on myofibroblast contractility of collagen gels[10], [11]. Importance of RhoA as a main regulatory factor was further supported by studies using three-dimensional myofibroblast-populated collagen gels[12], [13]. Moreover, using two-dimensional deformable culture substrates (PDMS substrate) that allow visualization of cell contractile forces, showed that inhibition of Rho could block myofibroblast contraction, whereas stimulation with lysophosphatic acid, an upstream effector of Rho, enhanced force development. In an interesting study contrary to aforementioned findings, it was shown that cytosolic calcium was the main regulator of three-dimensional collagen gel contraction. Additionally, interfering with Ca2+ signaling had a more dramatic effect on collagen contraction by myofibroblastic hepatic stellate cells than modulation of RhoA-dependent processes. Similar induction of myofibroblast contraction was obtained when treating two dimensional cell cultures with agonists that induce cytosolic Ca2+ transients.
In vitro findings on myofibroblast contraction mechanism have led to the development of an improved “lock-step” model of ECM remodeling. This model explains how global tissue remodeling can result from myofibroblasts contraction followed by ECM/substrate stabilization. The ECM stabilization is a result of secreted collagens and other ECM molecules by myofibroblasts themselves. In this process RhoA/ROCK signaling is responsible for promoting sustained and strong isometric cell contractions, which will generate small deformation in individual collagen fibrils. Such tension-released fibrils are then available to the weak and short-ranged microcontractions generated by periodic cytosolic calcium presence. The gradually raising tension in locally pulled fibrils will increasingly further local translocation. Next, the new fibril conformation has to be stabilized, possibly by local collagen digestion, deposition of new collagen fibrils, and crosslinking with the existent ECM[5], [14]. Although the details of this presumed remodeling step are not explored, it has been shown that collagen remodeling by matrix metalloproteinases can be regulated by mechanical tension. The new stabilized ECM environment can now sustain tissue stress and myofibroblasts are able to move around and spread out. These processes would result in irreversible contractures characteristic for fibrosis rather than reversible contraction [15].
On top of increasing the contractility through lock and key mechanism, myofibroblasts exhibit another unique cellular behavior that would result stiffening the soft compliant lung alveoli membrane and lung tissue. Myofibroblasts actively deposit proteins, mainly Collagen I, into extracellular matrix. When matrix deposition is coupled with induction of contractility, myofibroblasts start remodeling their environment. Moreover, it’s been very well documented that increase in tissue stiffness would result in higher fibroblast and myofibroblast activity. This could be considered as a positive feedback loop. In which the cells and environment constantly influence each other to the degree that the increase in stiffness is irreversible and the lung tissue is left with permanent fibrotic scar. This would decrease lung compliance and disrupts lung homeostasis. The question may arise regarding how this pathophysiological cycle is started? Recent studies have shown that besides triggering the contractility cascade as a result of fibroblast activity, multiple other mechanisms result in early changes in structure of ECM fibers and tissue stiffening. For instance, collagen cross-linking by lysyl oxidases takes place due to initial tissue stiffening, which leads to developing stable collagen cross-links by the activities of LOXL2 enzyme [16]. Additionally, myofibroblasts are present in abundance in areas of high ECM expression and represents the key source of intensified cytokine gene expression, release of inflammatory mediators, and epithelial injury, all of which are considered to be key factors 2 in preserving the cycle of injury and fibrosis. Another often overlooked result of increase in contractility is release of latent TGF-β. Which was produced by immune cells and myofibroblasts themselves. These secreted TGF-β later gets bound to ECM fibers and would be released as a result of application of α-SMA generated tensile forces. Where stress fibers act on transmembrane integrins, these integrins dock on latency-associated peptide (LAP) and latent TGF-β–binding protein-1 (LTBP-1) and by forcing conformational changes on them latent TGF-β is released from its pocket inside large latent complex (LLC) [3], [4], [7], [17].
Transformin Growth Factor (TGF-β)
TGF-β is the most potent profibrotic cytokine known, which as mentioned above in normal healthy tissue it’s mostly stored in the ECM inside LLC. The expression levels of TGF-β as a result of fibrosis is strongly increased, for instance there are well documented studies showing this elevation in several fibrotic lung diseases, including chronic airway remodeling in asthma, chronic obstructive pulmonary diseas, and interstitial pulmonary disease [18].
The TGF-β superfamily of growth factors consists of multiple proteins that have key roles in a wide range of physiological processes, such as stem cell pluripotency, cell fate determination, proliferation, and differentiation. Primarily, in lung fibrosis they are an important player in activation of myofibroblasts by enhancing expression of α-SMA stress fibers and collagen production. However, it also has positive and beneficial roles as well. TGF-β is known to control epithelial cell growth, inflammation, tissue homeostasis, and immune suppression in a variety of normal and pathologic adult tissues. Therefore making it difficult to battle lung fibrosis by just TGF-β inhibition treatments.
After TGF-β1 binds to the complex of TGF-β1 -receptor type I and II in the cell membrane. Cascade of signal propagation starts. SMAD2/3 phosphorylation gets initiated by receptors kinase activity. After phosphorylation SMAD 2/3 translocate into the nucleus and activates fibrogenic transcription factors. So much so that, in fibrosis, as the TGF-β1 levels are increased so does the nuclear accumulation of SMADs. However, studies have shown that SMAD inhibition does not slow down the fibrosis which implies that TGF-β1 binding to cellular membrane activates multiple transcriptional cascades in myofibroblsts.
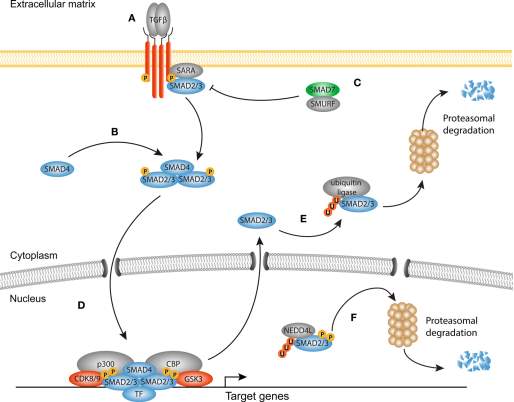
It has also become clearer that TGF-β1 signaling pathway works in harmony with WNT and YAP/TAZ canonical signaling. In-Vitro and animal studies have shown that in lung fibrosis there’s an increase in YAP/TAZ nuclear localization and the YAP/TAZ deficient fibroblasts are less responsive to TGF-β1 [19].
TGF-β, WNT, and YAP/TAZ output activity depends on nuclear translocation of its transcriptional factors. YAP/TAZ also regulates mechanical stress to accelerate fibrotic process. Transcription of α-SMA stress fiber and actin assembly are associated by nuclear translocation of YAP/TAZ and myocardin-related transcription factors (MRTFs). However their pathways are initiated by different response to tension. Nuclear accumulation of MRTFs is caused by release from G-actin, while TAZ/YAP activity is regulated by cytoskeletal tension. In in-Vitro studies on lung fibroblasts revealed that transcription factors changes as a result of substrate stiffness are the same as YAP/TAZ regulated ones. In lung fibrosis, with upregulated TGF-β expression, abnormal activation of TAZ/YAP may help elevating fibroblast activity. Overall, this cross pathway communication causes an increase in variety of phenotypes found in fibrotic disorders. After activating transcription factors act on chromatin and would boost or repress the target genes.
As a result of these activities in myofibroblasts, de novo expression of smooth muscle α-actin (αSMA) is identified [20]. Moreover, this coordinated communication makes myofibroblasts to make and secrete ECM components, such as collagens, glycoproteins, and proteoglycans, resulting in the formation of fibrous scar tissue. Further cross-linking of collagen by LOXL2 enzyme or cellular contractility makes it highly resistant to protease degradation [4], [20].
They also start to express the ED-A splice of cellular fibronectin. Organized and stable expression of ED-A Fibronectin at the plasma membrane Membrane is important in connecting actin and stress fibers to protruding integrin molecules bound to ECM. This is crucial in conversion of extracellular mechanical into biochemical cues that are transferred to the nucleus [21].
After this brief review of the importance of myfibroblast cells and TGF-β, as a fibrosis promoter cytokine, current and previous In-Vitro models for lung fibrosis will be discussed.
Lung fibrosis models
According to an NIH ARRA impact report, IPF is an untreatable fibrotic lung disease of unknown etiology. IPF is a fatal disease that is known to remodel the alveolar tissue, increase its stiffness (scarring) and impede oxygen transfer to the blood. Most patients survive only 2–5 years after diagnosis. The disease affects approximately 200,000 Americans and accounts for 40,000 deaths annually. In 2014, the U.S. Food and Drug Administration (FDA) approved two anti-fibrotic drugs, pirfenidone and nintedanib, for attenuation of functional decline in IPF. Pirfenidone has shown to reduce the inflammatory response in fibrosis and also reduce the decline in forced vital capacity (FVC). Nintedanib, on the other hand, targets several kinase pathway [6], [22], [23]. For instance, it targets vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF) and platelet derived growth factor (PDGF), reducing forced vital capacity (FVC) in patients with mild to moderate IPF disease. Besides these two pharmaceutical compounds, many other investigational new drugs (INDs) have failed in phase II/III of human clinical trials despite being successful in preclinical In-Vitro and animal studies. One of the major reasons for this inconsistency is the lack of functional and accurate animal and In-Vitro models that could accurately capture both pathological and mechanical aspects of lung fibrosis.
Various animal models have been developed over the years but each model is limited features of human IPF that they could imitate. Currently no animal model exists that can fully replicate all features of human IPF, especially the progressive and irreversible nature of the condition. Alternatively, there have been In vitro models of idiopathic lung fibrosis with human cells that could replicate key aspects of the disease otherwise not reproducible in animal models. Therefore, engineered in-vitro models would be a promising replacement or at least complement to imperfect animal models. In order to understand the necessity of In-Vitro models current animal models are briefly reviewed here.
Animal models
Several animal models have been developed to study the lung fibrosis. One of the main obstacles is that, the translation of results from animal models to humans trials, is complicated since animal used for the studies have different airway anatomy, lung cell biology and immunity. On top of that, strain based differences between animals and heterogeneity of disease development, make it more difficult to produce clinically relevant fibrosis models. Currently no animal model fully recapitulates the nature of lung fibrosis or the pathophysiology of usual interstitial pneumonia [24], [25].
The first developed and most used model is bleomycin model in rodents. When administered intratracheally, bleomycin causes inflammatory and fibrotic reactions within a short period of time (~1–2 weeks). This type of induced lung fibrosis features elevated cytokines related to fibrosis such as, interleukin-1 (IL-1), tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6) and interferon- γ (INF-γ), which results in an increase in pro-fibrotic markers including TGF- β1, fibronectin ED-A and procollagen-1. Fibrosis can be confirmed by day 14, with maximal responses noted at days 21–28 [26]. One of the main advantages of this model is simplicity and reproducibility. However, since the fibrosis markers and changes start to resolve after 28 days period, the major disadvantage of using bleomycin is the inability to model the chronic nature of IPF.
Another more sustained model is the fluorescein isothiocyanate (FITC) model of pulmonary fibrosis is like the bleomycin model in that inflammation precedes fibrosis formation. However, this model is not considered clinically relevant and dissimilarity in fibrotic responses have been observed the next model is fibrosis by the irradiation which is clinically relevant. With this model researchers can distinguish differences among inbred mouse strains, facilitating studies of genes involved in pulmonary fibrosis. However, it is too expensive [26].
In conclusion, although current animal models cannot recapitulate the inherent nature of IPF, they are still widely used to investigate signaling pathways and cellular interactions in fibrosis. Although recent advances have been made in creating models that study epithelial injury or epithelial-mesenchymal cross talk, ATS (American Thoracic Society) has recommended some necessary future works like intratracheal bleomycin administration in young male mice, using hydroxyproline measurements for collagen accumulation, alongside histologic assessments, for pre-clinical testing, and the expansion of humanized models of lung fibrosis [24–26].
Engineered In-Vitro cell and tissue models
Since idiopathic pulmonary fibrosis affects both bronchiolar and alveolar air spaces, multiple cells types are involved in IPF development and progression. Including but not limited to, large and small airway epithelial cells, endothelial, mesenchymal-fibroblast cells in alveoli membrane and immune cells. To this date most of existing understanding and knowledge on pathophysiology and pathways underlying IPF comes from studies performed on 2D cell cultures assays. To recapitulate the In-Vivo conditions primary and immortalized cell line have been used. Cell lines such as, BEAS-2B (B2B) large airway epithelial cells, non-small cell adenocarcinoma A549 and IMR-90 lung fibroblast cells are among the preferred choice of cell systems [27]. However, since immortalized cell line have been genetically manipulated and changes in their phenotype are permanent, they are not deemed as an accurate cellular model. Recent works have shifted their focus on using primary human lung cells.
In an in vivo environment cells are within a complex three-dimensional (3D) environment. In 3D connective tissue cell experience mechanical signals, extracellularmatrix (ECM) interactions, growth factor gradients and heterotypic interactions with mixed cell types. Cell in 3D express different integrin signaling and polarity compared, as a result 2D and 3D systems have different effects on modulating cellular phenotype.While 2D systems limit cellular migration and chemotaxis studies in the X and Y planes, 3D culture enable studies of cell mobility, orientation and cell-matrix interactions in all three (X, Y, Z) planes. Many researchers have categorized transwell, ECM and sandwich culture systems as 2.5D. In addition to movement physical dimension, it seems the difference in 2D and 2.5D arises from how the surrounding environment affects cell-cell interaction and organization. For instance, by contrast to culturing on plastic dish, culturing cells on top of ECM hydrogel helps formation of higher level 3D epithelial acinar structures are formed. Lung fibroblast cells are in the focus of tissue stiffening and remodeling in IPF and these cells are under influence of biochemical and biomechanical signals. Therefore, variety of 2.5D and 3D systems were studied. In contrary to 2D systems where fibroblasts exhibit forced apical-basal polarity, fibroblasts cultured in 3D systems experience no forced polarity, resembling physiological conditions. Plus, compared to 2D culture, 3D environment modulates different mechanotransduction signaling profile with integrin adhesions and cellular traction in all three planes. Here, previous culture systems from 2D to 2.5 and 3D are discussed[28], [29].
Two dimensional culture studies
Tissue culture plastic (TCP) and glass substrates provide researchers with a standard platform to evaluate single cell responses to soluble growth factor and extracellular matrix cues. Although culture systems are becoming more and more sophisticated, the testing of single molecular effects on individual cells without neighboring cells/molecules are still relevant as they provides important information. Another advantage of using 2D cultures are the suitability of this method for profiling cells for profibrotic and fibrosis supressing gene expression following different treatments. Additionally, these 2D culture platforms are often used in conjunction with animal studies or other culture types. For example, bleomycin animal studies identifying novel proteins have used 2D culture substrates to further elaborate single protein effects on lung cells. Alongside TCP and glass substrates, mechanically tunable substrates have been used to evaluate the effect of substrate stiffness on lung fibroblasts and myofibroblasts. Mechanically tunable polydimethylsiloxane (PDMS) substrates and polyacrylamide (PA) hydrogels have frequently been used by researchers to study influence of various mechanical environment on lung pulmonary fibrosis. PA gels are prepared by mixing different ratios of acrylamide and bis-acrylamide (cross linking agent). Following polymerization, ECM proteins, collagen type I, are covalently crosslinked to the surface of the gel and will promote cellular attachment.
After the publishied study by Engler et al.[30], which shows the how to fabricate mechanically tuned PA hydrogel, researchers have studied effect of different substrate stiffness on myofibroblasts. The stiffness of the substrate is based on lung tissue youngs modulus in healthy and diseased conditions. Balestrini et al. showed that myofibroblasts cultured on stiff substrates possess ‘mechanical memory’[4]. Using mechanically tuned PDMS substrates with stiffnesses between 5 to 100 kPa, the authors showed increased myofibroblast differentiation accompanied by increased alpha-smooth muscle actin (α-SMA) expression. Which was significantly obvious on stiff 100 kPa substrates after 1 week of culture compared to soft 5 kPa substrates. The expression was sustained even when the fibroblasts previously cultured on 100kPa substrate for three weeks, were transferred onto the soft 5 kPa substrate. In another work Markinovic et al. employed mechanically tuned PA hydrogels and showed that fibroblasts isolated from patients with IPF are phenotypically different from normal fibroblasts. Interestingly fibrotic tissue derived fibroblasts exhibited increase in proliferation with increase in matrix stiffness. However, when compared to fibroblast derived from healthy tissues, IPF fibroblasts possessed significantly reduced proliferative activity [31]. Furthermore, it was shown by culturing in soft environment, distinctive contractility of IPF fibroblasts is significantly reduced.
Despite their commonly application in cellular assays, the 2D culture models on plastic and glass substrates have serious limitation. First and foremost, the 2D models are unable to mimic the 3D nature of lung tissue. Tissue culture plastic and glass are extremely stiff (~1 GPa) substrates which affects cellular phenotype, and stimulates genetic expressions that are not associated with native cellular conditions. Moreover, standard culture substrates do not allow studies evaluating cellular mechanics and tissue remodeling due to lack of ECM. Mechanically tunable substrates can replicate normal and diseased lung mechanics, however all 2D substrates force fibroblasts to attain an apical-basal polarity, unlike their natural state in vivo, where fibroblasts exhibit a more rounded morphology with integrin cell adhesions in all three planes.
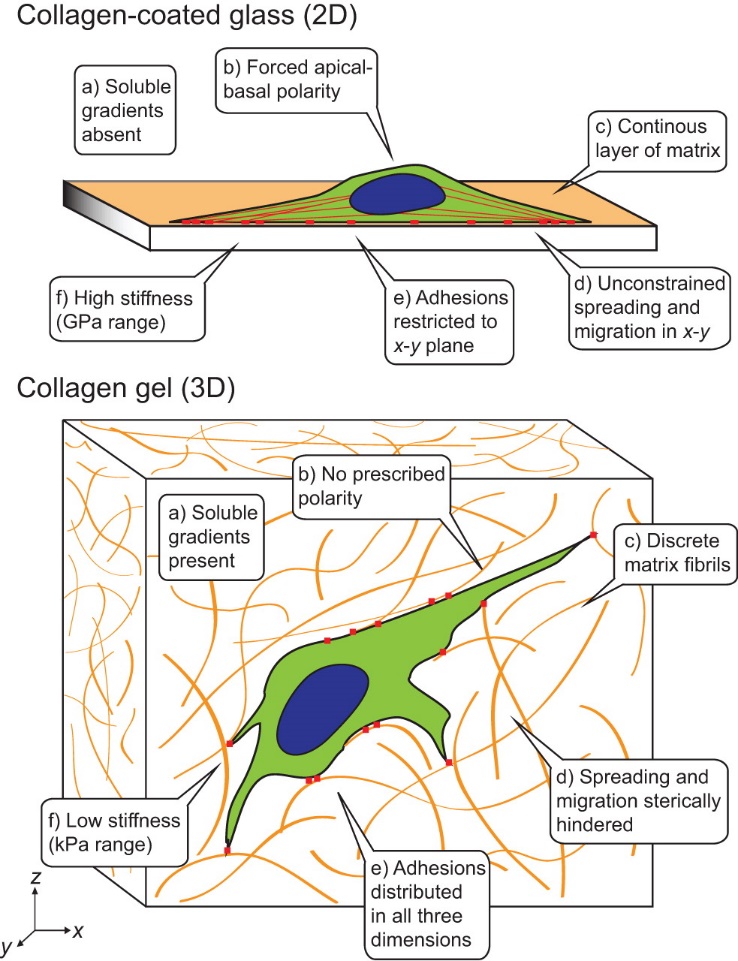
Further, 2D culture substrates cannot model exposure to gradient soluble cues found in vivo and the variety of cell-cell, cell-matrix interactions experienced by cells within the normal human lung. When it comes to lung airway modeling which requires epithelial cells, it is necessary for the In-Vitro models to function under air liquid interface (ALI). 2D substrates fall short on promoting ALI culture required for basal cell differentiation and airway maturation. Hence, 2D studies of epithelial cells are relatively short-term studies, and not representative of in vivo lung epithelia.
One of the commonly used techniques for ALI is transwell culture. In fibrosis, epithelial injury comes before fibroblast activation, therefore transwell systems have been employed extensively for co-culture studies with epithelial cells. Epithelial ALI co-cultures are often done with lung microvascular endothelial cells and/or fibroblasts. Epithelial-fibroblast co-cultures have been studied for the purpose of investigating pathological paracrine signaling. This signaling mechanism has been connected to myofibroblasts activation. Epa et al. utilized transwell systems to reveal a new role for anti-fibrotic prostaglandin-E2 (PGE2) signaling observed in human IPF patients [32]. Which showed epithelial cells from PGE2, inhibited TGF induction of fibroblasts in paracrine way. Other study co-cultured non-small adenocarcinoma cells (A549) with healthy and IPF derived fibroblasts. That study revealed the importance of paracrine PDGF signaling in fibroblast migration. The transwell method has been really helpful in shedding lights on paracrine activity of epithelial and endothelial cells on lung fibroblasts.
One other method in better modeling lung fibrosis in 2D cultures are culturing the cells on layer of proteins expresses or constitute basement membrane in lung tissue. The basement membrane plays a key role not only in chemical signaling but also in providing mechanical cues and structural integrity. For instance, airway epithelial cells and microvascular endothelial cells laid out on top of underlying basement membrane, possess polarized assembly. Studies have shown that airway epithelial and microvascular endothelial cells cultured on basement membrane such as Matrigel make polarized and complex structures, not reproducible by 2D systems. In one of the studies, Yu et al. showed that human alveolar type II cells (ATII) seeded on top of Matrigel produce hollow cysts lined by a polarized monolayer of cells. That study also revealed the formed structure is also capable of producing surfactant. Matrigel is a reconstituted basement membrane extract from the Engelbreth-HolmSwarm (EHS) murine tumor. This protein is a great source of laminin, growth factors, entactin/nidogen, type IV collagen and heparan sulfate proteoglycan.
Basement membrane cultures was also practiced using microvascular lung endothelial cells as an In-Vitro platform to study of anti-fibrotic agents. Much like epithelial cells, these cells also underwent 3D remodeling when cultured on Matrigel membrane. Where, they formed 3D tubular structures mimicking angiogenic programming. Evrard et al. cultured endothelial progenitor cells on Matrigel membranes and showed that TGF-β1 stimulated proangiogenic signaling by increasing cell viability and migration, but did not affect capillary network formation [33]. Cosgrove et al. also used Matrigel assay to investigate the profibrotic effect of IPF lung homogenates on endothelial tubulogenesis.
Sandwich culture is another attempt studied by the researchers to bring 2D culture closer to modeling native tissues. In this method cells are culture between two medium usually fabricated with hydrogels. First the cells are plated on top of the substrate hydrogel or culture plate, then another layer of gel is casted on top of the plated cells. The sandwich culture has two important advantageous over conventional 2D culture on plastic dish. One is the option for exposing cells to two layer of tunable mechanical substrate, and the second is ability to support integrin adhesion in all three directions of X,Y and Z; similar to 3D environment. Other studies have used this platform for exploring cell-cell and cell-ECM interactions, however, few studies have adopted this technique for pulmonary fibrosis investigations. Additionally, sandwich cultures have the potential to replicate matrix stiffness gradients like the environment in IPF fibroblastic foci, where fibroblasts could be grown between substrates of different stiffness, simulating normal and fibrotic status of the disease. Studying cell migration and chemotaxis is also feasible if right substrates are used.
Considering the aforementioned adjustments to conventional 2D culture systems, these models still hold challenging limitations. Some common limitations, previously mentioned for 2D cultures, are observed for transwell and basement membrane models. As they cannot provide soluble growth factor gradient and 3D integrin adhesion to surrounding. Lack of mechanical tenability is another shared concern. Mechanical tenability is one the less explored aspects of culture systems. However, when it comes to In-Vitro modeling of a disease that target soft tissues, such as fibrosis, this aspect is an essential key to create more accurate disease models. Sandwich models have tried to overcome these challenges by providing integrin adhesion in all direction, exposure to tunable substrates and gradient growth factor. However, choice of the hydrogel used in these systems directly affects the ability of sandwich cultures to enable 3D migration, chemotaxis studies. Despite, enabling more enhanced cell-cell and cell-ECM interactions, sandwich cultures cannot support dynamic tissue remodeling and cell matrix interaction that exists in lung tissue.
Three dimensional culture models
In order for 3D model to be successful and overcome 2D culture limitations it has to efficiently replicate cell-cell and cell-matrix interactions, while mimicking native matrix stiffness and micro/macro structure. A functional model should promote migration and movement, chemotaxis, and cellular traction. The environment should favor cell adhesions, in all three planes. Further, these systems should have potential to replicate soluble growth factor gradients and support ALI epithelial maturation, comparable to native lung conditions.
Here, we would discuss the current three dimensional in vitro culture systems that were investigated to create native lung tissue microenvironment with greater accuracy compared to mentioned 2D models. Various researchers have proposed different 3D models to replicate paracrine cellular and matrix signaling that drive fibrotic lung disease progression. Some of these systems include, hydrogels, decullularized matrices, spheroids and organoids and lab on chip.
Hydrogel culture systems
Hydrogels are water-swollen cross-linked natural or synthetic polymers. By modifying chains network or crosslinking mechanism, hydrogels could be mechanically tuned to replicate mechanical moduli of lung tissue. The composition of the hydrogel can also be tailored to match human tissue composition. Hydrogels fabricated from collagen type I are the most-studied 3D culture system for modeling pulmonary fibrosis. This is due to the fact that collagen type I is the major ECM found in lung membranous tissue and also within the fibroblastic foci in IPF. Acid-solubilized collagen I, when neutralized, undergoes fibrillogenesis which creates entangled collagen fibers. By adding fibroblasts during polymerization; cells would be encapsulated and attached to collagen fibers. Since the fibroblast’s integrin are adhered in all the directions, cells start to exert force and contract the collagen hydrogel network. Either, floating or anchored matrices has been used to model early and late stages of wound remodeling. Other cell types including epithelial and endothelial cells also have been shown to contract collagen matrices. Arora et al. performed 3Dculture of fibroblasts in floating and plated collagen and studies their gel contraction under TGF-β1 stimulus. They showed that by adding the TGF-β1, expressions of α-SMA was elevated which resulted in proportional change in gel compliance. Floating collagen gels with the least compliance had the least expression of α-SMA. In comparison, moderately compliant anchored gels and stiff collagen-coated plastic had moderate and high levels of α-SMA and integrin (α1, β1) expression, respectively. Although interesting, his findings does not match the real effect of α-SMA and fibroblast activation on lung tissue. Since areas of stiffened scar tissue that has lower compliance; has higher expression of α-SMA. Other researchers have also used the 3D fibroblast culture in collagen to study influence of other important cytokines present in IPF such as platelet derived growth factor or PDGF. Peng et al. studied role of PDGF in collagen contraction assay. Their work revealed that different concentrations of PDGF stimulates gel contraction in restrained matrices. They also tested lysophosphatidic acid (LPA) which showed the same effect as PDGF. Among other natural hydrogels, fibrin and hyaluronic hydrogels have been considered for fibrotic disease modeling, the latter is overexpressed in fibrotic scars in human IPF. One of the significant limitations of natural hydrogels is their lack of tenability in of mechanical properties. Especially, when it’s needed to match the stiffness of fibrotic tissue. Another restriction of these natural hydrogel is their culture stability due to degradation and contraction. To address this challenge, other natural polymers have been used in composition with collagen. For example, mixture of collagen and natural silk fibroin hydrogels have been investigated. Mixing silk with collagen type I generated hybrid silk-collagen-type-I tyrosine crosslinked hydrogel systems for 3D culture of fibroblasts, where the silk protein controls hydrogel modulus and collagen type I supports cellular integrin adhesions. The silk-collagen composite hydrogel can provide mechanical moduli close to properties of human lung tissue. Fibroblast in 3D culture inside this composite have exhibited reduces gel contraction and longer culture stability. Researchers have proved the feasibility of myofibroblast activation and IPF modeling using the silk-collagen type I hydrogel. Although not desirable, synthetic polymers also have been used for In-Vitro 3D modeling of IPF. Most notably, polyacrylamide hydrogels (PA) and poly-ethylene-glycol (PEG) hydrogels, have been used to study lung fibrosis. The main advantage is accuracy in mechanical tenability of these synthetic hydrogels; however, toxicity and lack of biocompatible environment for cell adhesion, spreading and tissue remodeling are among top reasons that these synthetic hydrogels are not the most ideal option to replicate lung tissue compared to natural hydrogels.
Decellularized human and animal matrices
Decellularized matrices are derived by removing the cells from the tissue or organ. This is achieved by using detergents, leaving a complex mixture of structure that retains all the key proteins that constitute the extra cellular matrix. The method used to apply the detergent and the duration of the process, affects the loss or preserving the ECM proteins. One of the main aspects of decellularization is their application for in vivo implantation, where the other utilization is performing studies of matrix-related cell interactions, as decellularized matrices could be accepted as most ideal 3D matrices for in vitro study of pulmonary fibrosis. They retain native 3D structure and ECM protein composition. The protocols are designed to eliminate any potentials of later immunological reactions by removing cellular and nuclear materials however, during the process some of the elastin, collagen and glycosaminoglycans (GAGs) are also damaged or removed. Therefore, if the matrix is intended to be employed as an In-Vitro IPF model, determining the ideal detergent protocol in order to maintain the native stiffness and avoid loss of ECM proteins is critical. For example, Past studies using peracetic acid have confirmed acceptable preservation of VEGF, basic-FGF and TGF-β in decellularized matrices. Besides human tissues, decellularized matrix could also be fibroblast derived. Fibroblast derived matrices are economical, quick and rich with ECM latent growth factors. This matrix is processed after 3-4 weeks of fibroblast culture. Some researchers have used this matrix to provide more structurally accurate environment that could secrete TGF-β1 latent complex within ECM. Klingberg et al. used fibroblast derived matrices to investigate activation mechanism and secretion profile of TGF-β1 from the ECM. By comparing cell-derived matrices from fibroblasts and myofibroblasts, the authors showed that myofibroblast-derived matrices contained relatively higher amount of TGF-β1. Some of the limitations of fibroblast derived matrices stems from the matrix microenvironment and composition that are produced by these cells, compared to heterogeneous ECM composition, complex structure of native lung tissue. Moreover, for the most part each matrix produced by fibroblast cultures need to be characterized for the precise ECM composition. Providing tunable mechanical environment with stiffness moduli matching IPF lung is another issue. Lastly, the thickness of the generated matrix is limited, although they are considered 3D structures.
Unlike fibroblast derived matrices, decellularized lungs have developed to be an ideal and accurate model for in vitro modeling of IPF. Acellular lung tissues have the correct ECM composition of the native lung, almost identical structure and stiffness. More importantly, not only it can be obtained from healthy, normal lung but also, from human IPF donors. This factor allows the investigation of profibrotic phenotypes induced by pathologically-modified ECM and matrix stiffness. Booth et al. established the use of decellularized IPF human lungs as an in vitro model from normal or IPF donors. The lung tissues were processed by perfusion using non-ionic TritionX-100 and ionic detergent sodium deoxycholate to eliminate nuclear and cellular material. The investigators tested decellularized pieces by seeding them with normal lung fibroblasts. Normal fibroblasts seeded within IPF matrices, differentiated into myofibroblasts, and expressed increased α-SMA positive stress fibers expression; compared to cells seeded in normal lung matrices.
On the other hand, they did not observe any difference in TGF-β1 bioactivity between normal and IPF matrices. Therefore, ECM induced myofibroblastic phenotype was concluded to be as a result of TGF-β1 independent mechanisms. In another study, Parker et al. utilized IPF derived lung matrices in a microarray analysis, which led to identification of miR-29 role in lung fibrosis.
One the main limitations of employing human lung tissue for researches; comes from their availability, plus, acquiring diseased IPF tissue is even more challenging due to the rare nature of the disease. Further, a limiting human tissue supply can impede high throughput studies and the capacity of these systems to support anti-fibrotic drug testing. Another important questions is regarding cellular interaction with the matrix, for instance, myofibroblasts are often found within fibroblastic foci which is rich with collagen I, however when fibroblasts are reseeded within acellular matrices, they are subjected to ECM cues which are not anatomically representative of fibroblastic foci microenvironment. This is due to heterogeneity of lung tissue in different regions as confirmed by quantitative proteomic analysis of ECM of the human lung. The impact of ECM heterogeneity within lungs on investigation of profibrotic cellular phenotype is unclear.
Lung organoids and spheroids
Lung organoids and spheroids culture can enable cell-cell, cell-matrix interactions, structure and functionality that are missing from other 3D culture systems. Although sounds similar, there are differences between organoids and spheroids. Lung spheroids come from to adult lung cells or lung stem cell cultured in the form of aggregates utilizing low-adhesion plates, hanging drop cultures, suspension cultures or micropatterned plates. Lung organoids are more complexed. Organoids are self-assembling structures of lung stem cells that replicates series lung developmental process, with discrete pseudoglandular, canalicular and saccular stages. Therefore with limitations of In vitro culture systems, organoids experience incomplete development, for example, maturation toward the alveolar stage with septa formation and microvascular maturation; are not feasible. Henry et al. fabricated lung spheroids from culturing adult lung cells in suspension. These suspended spheroids were positively expressing pro-surfactant protein-C (pro-SPC). Next, these lung spheroid cells were cultured on Matrigel in order to encourage self-assembly. Through self-assembly motion the spheroids turned into alveoli like structures with positive immunostaining for aquaporin, indicating mature lung epithelial cell differentiation. The authors further investigated therapeutic potentials of these spheroids in pulmonary fibrosis. Therefore, lung spheroid cells were injected via tail vein of immunodeficient mice 24 hour post-bleomycin administration. Hematoxylin & eosin (H&E) staining showed parenchymal infiltration of these lung spheroid cells and importantly they significantly inhibited bleomycin-induced fibrosis. The authors showed that fibrotic scars were lower in spheroid treated groups even compared to the mice that were injected with mesenchymal stem cells. They did not determine the underlying mechanism, but it was hypothesized that spheroid cells promoted regeneration through indirect paracrine mechanisms, similar to protective mechanisms observed with MSC treatments within myocardial infarcts.
In order to create organoids, induced pluripotent stem cells (iPSC) were used by Chen et al. they utilized iPSCs to model lung fibrosis as a result of viral infections. The differentiation protocol needed 6 months of organoid 3D culture inside Matrigel. They showed the organoids differentiation went along specific lung developmental stages, up to the saccular phase, which takes place in second trimester of embryonic development. Next, the organoids were either infected with virus or underwent gene deletion by CRIPR method to model respiratory syncytial virus (RSV), Hermansky-Pudlak Syndrome (HPS) diseases respectively. During infection modeling the epithelial layer got swollen and started the shedding process as it was confirmed by confocal microscopy. The authors investigated the lung organoid cultures and found increased number of mesenchymal cells and expression of mesenchymal markers including PDGF-receptor-A, PDGF-receptor- B, α-SMA, vimentin and cluster of differentiation-90 (CD90). The authors also noticed an increased presence of profibrotic ECM proteins including collagen types I and III and fibronectin. Similar to HPS syndrome, mutant iPSC derived epithelial cells within lung organoids drove abnormal gene expression and ECM production in mesenchymal cells. The main advantage of the spheroid and organoids are their ability to create higher organ level 3D complexity and functionality compared to hydrogel, tissue derived matrices and conventional 2D culture system. However, their complicated differentiation protocol comes with the cost of being low throughput and inconsistent. This is even more significant when it comes to In-Vitro screening of drug. To address this challenge, currently improvements are being made to enable spheroid cultures in low adhesion high density microplate formats including 384-well and 1536-well, combined with automation systems. In terms of consistency, it remain the main limitations of these models, highly variable size and shape of these spheroids/organoids make a challenging quest to acquire biologically meaningful and reliable results, especially during compound screening. Imaging difficulties due to cellular migration and their complex 3D nature, is another familiar challenges of these structures.
Organ on chip
Organ on chip devices are bioengineered devices which summarize physiological phenomenon or model organ/tissue structure in miniature chips created using lithography techniques that could sometimes serve as a high throughput system in static or dynamic culture. One of the main features of creating organ on chip rather than reductionist aspect of these devices, are enabling researchers to have exercise precise control over the features and pre-set conditions which makes the model more accurate and reliable.
Felder et al. fabricated a microfluidic wound healing assay to study effect of gastric refluxes acid on alveolar tissue injury, a significant etiological factor in human IPF. The same design was employed later, with normal lung fibroblasts to study influence of breathing in form of cyclic stretching strain on the cells activation toward myofibroblasts. The results showed that cyclic strain reduced TGF-β1 induced myofibroblast markers including α-SMA, collagen types I connective tissue growth factor. In another study done be Sellgren et al. multi-layer microfluidic system was studied for co culturing fibroblasts with endothelial or epithelial cells which were separated by porous membrane. This device was a modified version of the original lung on chip micro device fabricated by Huh et al. their lung on chip system was used to co-culture monolayers of endothelial and epithelial cells which allowed investigating outcome of shear and strain caused by breathing and perfusion. One of the disadvantages of lung on chip system was inability to quantify mechanical outcome as a result of fibrosis progression. Although the authors tried to utilize their device to model lung infection, some critics noted to similarities between their method and transwell culture systems. Therefore, Fibrosis-on-chip devices have been fabricated by combining layer of hydrogel, in order to constitute a three-dimensional (3D) system. Mondrinos et al. were one of the groups that employed that method and encapsulated fibroblasts within collagen hydrogels inside the lung on chip device, to replicate fibrosis on chip. Through adding growth factors and media optimization, they were able to activate fibroblasts, similar to pathological fibroblastic foci. Next, they successfully amplified fibrosis by perfusion of monocyte derived macrophages and cytokines promoting M2 macrophage phenotype. Despite, their effort to create a fibrosis model on a chip, anti-fibrotic drug testing by identifying signaling pathways could not be achieved on their model. This drawback could be due to inherent limitations, including technical difficulties like bubble formation which are known to injure cells in these devices, contraction/degradation of ECM gels or coatings. On top of that, absence of mechanical tenability and failing to mimic stiffness associated with lung fibrosis, overshadowed the successful features of their system. Regardless of these shortcomings, organ on chip devices have started to offer an exciting platform for researchers to model physiological and pathological actions in a high throughput fashion and close to an organ level complexity.
Cite This Work
To export a reference to this article please select a referencing style below:
Related Content
All TagsContent relating to: "chronic illness"
Chronic illnesses are non-communicable diseases which develop gradually over time. They generally cannot be solved instinctively and are barely to be cured ultimately. In most cases, chronic illnesses are more likely to be insidious where the patients have mild or no symptoms.
Related Articles